Summary
The family Rissoidae is a family of small-sized prosobranch, gastropods found all around the globe. They can be found in marine and brackish environments, with distribution ranging from the tropics to the Arctic and Antarctic (Ponder, 1985). Most Rissoids are found in high abundance through inter-tidal and shallow, sub-littoral environments feeding on algae (Avila, et al. 2012).There are however, a few species that have been discovered at continental shelves and in abyssal zones (Ponder, 1985). Species from the family Rissoidae, are quite difficult to identify due to their small size and similar physiology (Ponder, 1985). The unidentified species of this study shows similarities to a number of genus', such as Voorwindia and Setia. This study looks at physical, behavioural and functional characteristics of the unknown species to get a better understanding of its nature. Experiments were set up to test the movement behaviour and reproductive development of the unknown species.
Physical Description
Shell:
The Family Rissoidae is a group of small gastropods found in marine to brackish waters all around the world (Avila, et al. 2012; Ponder, 1985). Rissoid shells vary from smooth to sculptured, ovate to elongate-conical and thick to thin (Ponder, 1985). The shell is lacking a chitinous inner layer and often has a dotted or spiralled protoconch pattern (Ponder, 1985). The individual at the centre of this study demonstrates a smooth, partially transparent, elongate-conical shell; with a dashed,spiralling protoconch pattern (Figure 1).
 Figure 1: Shell of the Rissoidae species collected
Figure 1: Shell of the Rissoidae species collected
Body:
Measurements from ten individuals determined that adults had an average length of 2.3mm and width of 1.6mm. This matches sizes of the genus Voorwindia, which are characteristic of lengths up to 2.6mm (Ponder, 1985)
As with all prosobranchs, Rissoidae are completely torted, have asymmetrical visceral mass,a well developed head and two cephalic tentacles, with a lateral eye at the base of each (Ponder, 1985; Ruppert,et al. 2004). The muscular foot can vary from short to long but is always equipped with an anterior pedal gland (Ponder,1985).The undefined individuals were observed gliding upside down on the water’s surface. This is found in a number of Rissoidae members and is associated with the presence of a posterior pedal gland which allows individuals to string mucus off the water surface (Ponder,1985).
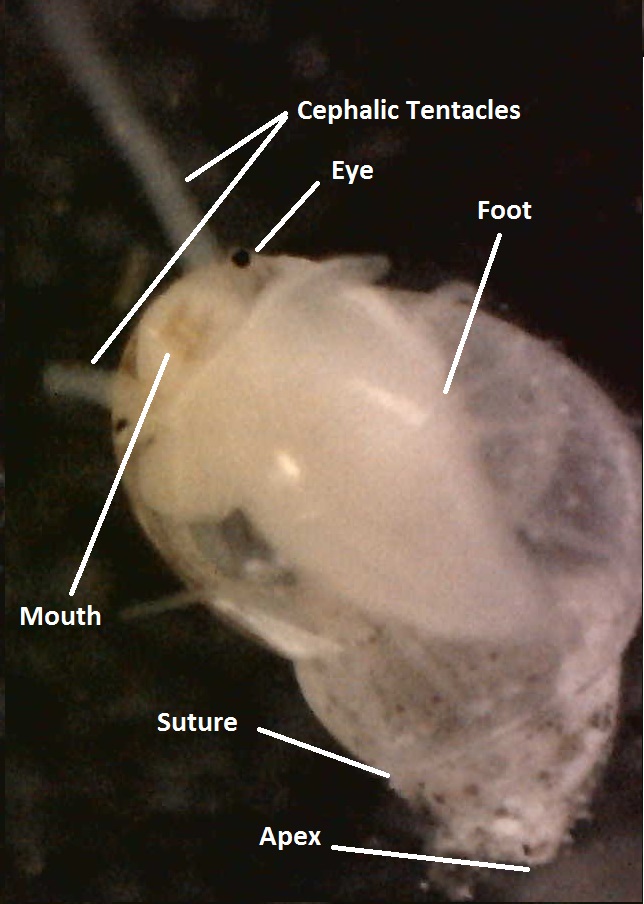
Figure 2: Key Physical features labelled on the Rissoidae individual collected. Operculum:
The operculum, visible in a number of gastropod taxa, is a small ‘lid’ that is retracted over the shell when the individual retracts (Ruppert,et al. 2004). This cap operates as a wall for individuals, protecting them from predators. The operculum of species from the family Rissoidae is rough, oval shaped and may vary in thickness (Ponder, 1985). An operculum was observed for the Rissoid species, fitting the thin, oval characteristics of the genus Voorwindia (Ponder, 1985).
Radula:
The radula is a flexible tongue-like structure,with transverse rows of small chitinious teeth (Ruppert,et al. 2004). This is a characteristic feature of mollusca, only absent in bivalves. Members of the Rissoidae can be identified by their taenioglossate radula. This is a radula which generally has seven teeth in each transverse row, with a layout of 3-1-3 (Figure 3). The central teeth can often be used to separate genera within Rissoidae (Ponder, 1985). The number of paired cusps, sometimes referred to as basal denticles, is the defining character (Ponder, 1985). Most genera only show one pair of cusps; however this does vary in other genera (Ponder, 1985).
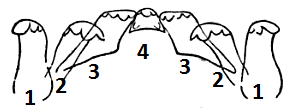
Figure 3: Taenioglossate Radula of Rissoidae family. Image modeled off illustration from Ruppert, et al. (2004)
Biogeographic Distribution
Members from the family Rissoidae have been found in seas all around the world, ranging from polar to temperate latitudes (Ponder, 1985).Genera such as Alvania, which have planktotrophic larvae, have extensive distributions that are associated with oceanic circulation (Ponder, 1985). Other genera, such as Attenuata, have direct development which tends to limit the organisms to a smaller geographic distribution (Ponder, 1985). This is argued in a paper by Avila (2013), who states that organisms with direct development may have the ability to raft. This may be achieved by algae breaking loose while the species attached (Ponder, 1985).
The distribution of two genera, that have similar morphological characteristics to the unidentified Rissoidae, were found on the Encyclopedia of Life. The distribution of the first genera, Voorwindia, is shown in Figure 4 below. This figure shows that Voorwindia has high distribution on the East coast of Australia, with some individuals also found around Japan. Figure 5 shows the distribution of the genus Setia has been found in seas north of Africa, in high abundances. From the distributions portrayed by the Encyclopedia of Life it would suggest that the unknown Rissoidae is more likely to be from the genus Voorwindia than Setia.
Local Distribution
Members of the Rissoidae are mostly found in inter-tidal and shallow littoral habitats feeding on algae (Ponder, 1985). Some inter-tidal species have been shown to live under rocks and feed on foraminifera (Ponder, 1985). There are also a number of abyssal species that have been discovered and this number is expected to increase as sampling methods advance (Ponder, 1985). The individuals used in this study are of an unknown origin. It is believed they were introduced to the tank with the gravel or on the marine plant which is abundant in the tank. The marine plant belongs to the genus Caulopa, which is a major group of seaweeds.
Species from the genus Voorwindia have been discovered off the south-east coast of Australia and shows similar characteristics to the individuals of this study (This is discussed under Evolution and Systematics). Voorwindia umblicata is found in estuaries and sheltered bays on the northern half of the New South Wales coast where it is associated with seagrass (Ponder, 1985). Sampled depths varied between 0-11 meters. As shown in the biogeographic tab, this is not the case for Setia, which is located predominantly in the northern Hemisphere (Avila, 2013; Ponder, 1985).
Locomotion & Movement
The muscular foot is the primary mechanism of locomotion for Gastropods; as well as functioning in prey capture, defence and reproduction in some species (Ruppert,et al. 2004). The shape, thickness and functionality of the foot may vary significantly between different gastropod species. The gastropod foot consists of a series of muscles and connective tissues that function in different movements. The columellar and tarsos are the two major muscles used for moving the gastropoda foot. The columellar muscle is responsible for the retraction of the individuals head and foot into the shell, as well as the twisting of the foot in regards to its shell. The columellar muscle is located dorsally in the foot’s center and its muscle tissue often acts as a hydrostatic skeleton for gastropods. The tarsos muscle is composed of fine muscle bundles and located on the foot sole. This muscle is responsible for locomotion in gastropods and may also function in other roles depending on the species.
The unknown Rissoid species, as mentioned has a posterior and anterior pedal gland structures on its foot. These glands specialize in the production of a mucus trail for the organism to glide over. The sole of many Rissoidae are equipped with pedal cilia which generate movement using the mucus layer provided by the pedal discs.
Behaviour experiment:
This experiment aims to investigate the behaviour of the unknown Risoid species in regards to movement and determine which sensory systems, if any, are being used to make these decisions (e.g. Mechanoreceptors, chemoreceptors and photoreceptors).
Question 1: Do they act as individuals or form clusters to better defend against predators and increase mating opportunities?
Method:
Eight individuals were collected from the downstairs aquarium and held in a large petrie dish with plant matter. Four individuals were spaced evenly apart in a 4cm petrie dish, see figure 6. Individuals were provided a number from 1-4 and their initial moves were noted. Observations were continued until the individuals had stopped moving to make sure they were in their preferred location. This was repeated five times to show any patterns in movement behaviour. Before each replicate the filtered water was replaced.
 Figure 6: Shows the four unidentified Rissoidae individuals shortly after initiating the first experiment (cluster test)
Figure 6: Shows the four unidentified Rissoidae individuals shortly after initiating the first experiment (cluster test)
Hypothesis: It is expected that individuals will move together and cluster to better improve their chance of survival and finding a mate.
Results:
It was found that the initial movement of all individuals, for the five replicates, was to the edge of the dish. However, once on the dishes edge, individuals would follow it around to other individuals on most occasions. The table below shows the percentage of individuals from each test that clustered after reaching the dish edge:
|
|
1
|
2
|
3
|
4
|
5
|
Average
|
|
Clustered (%)
|
100
|
50
|
50
|
75
|
75
|
70
|
|
Separate (%)
|
0
|
50
|
50
|
25
|
25
|
30
|
Table 1: Shows the percentage of individuals that clustered vs. stayed seperate over five replicates.
Once on the edge of the dish there is evidence that clustering becomes a factor. However, two replicates showed that only half of the individuals decided to cluster.
Discussion:
There was a clear trend of movement away from the petrie dish centre towards the edges in all five replicates. With no cover to hide under, individuals are completely exposed in the plates centre, leaving them vulnerable. However, by moving to the edge of the petrie dish they receive protection on one side.
Once on the side of the petrie dish, individuals could find each other and form a cluster of remain alone. On average a higher percentage of individuals were found to cluster together instead of stay alone.This supports the hypothesis that the unknown Risoid species clusters together. This may be done to better protect themselves or increase mating opportunities. However, a number of individuals were seen to prefer space. This could be associated with the competition for food and other resources when clustered together as a group. This suggests that the directional and interactive behaviour of the unknown Risoid species may be affected by whether individuals feel threatened, are searching for resources or have received reproductive stimulus.
Question 2: Do the unknown Risoid species demonstrate directional movement in the form of chemoreception and/or photoreception? Or is their movement relatively non-directional?
Method:
The same eight individuals were used from the first test, with 4cm petrie dishes, filtered seawater and four individuals per replicate. Three different tests were done to show the movement behaviour associated with different objects. Plant matter from the downstairs aquarium was used as an object that would demonstrate chemosensory. (Note: the marine plant was found in high abundance in the same tank as the unknown Risoid species population). Individual tests were also done with a clear and coloured piece of plastic to show if movement was associated with photoreception (Clean plastic). The initial movements were recorded (movement to object or dish edge). Observations were continued to confirm that movement was towards their preferred location. All tests were replicated five times with fresh filtered seawater to show patterns in behaviour.
Hypothesis: A higher percentage of movement towards the visible plastic when compared to the clear plastic will demonstrate photoreception; while chemoreception is evident from the movement towards the leaf/other individuals when compared to the plastics
Results:
The initial movements observed by individuals showed that objects did make a difference to the behaviour seen in experiment 1. There was evidence of movement towards the plant and coloured plastic over the five replicates, while, the clear plastic was shown relatively little interest. A percentage was calculated for each test replicate to show the percentage of individuals that moved towards the object (plant, clear or coloured plastic) rather than the petrie dish edge.
|
|
1
|
2
|
3
|
4
|
5
|
Average
|
|
Plant (%)
|
50
|
50
|
75
|
50
|
25
|
50
|
|
Clear Plastic (%)
|
0
|
0
|
0
|
25
|
0
|
5
|
|
Colour Plastic (%)
|
50
|
50
|
25
|
25
|
0
|
30
|
Table 2: Shows the percentage of individuals that moved to the object (Plant, clear plastic or colour plastic)
 Figure 7: Shows the four unidentified Rissoidae individuals in the coloured plastic test shortly after starting.
Figure 7: Shows the four unidentified Rissoidae individuals in the coloured plastic test shortly after starting.
 Figure 8: Shows the result from a plant matter test with two Rissoidae individuals on the object and two at the petrie dish edges.
Figure 8: Shows the result from a plant matter test with two Rissoidae individuals on the object and two at the petrie dish edges.
The low recognition of the clear plastic resulted in all individuals moving straight for the petrie dish edge, once there they move around as seen in the first experiment with some clustering and some staying alone. Now and then an individual would come across the clear plastic and check it out before moving on.
Discussion:
The tentacles of the unknown Risoid species demonstrated use as mechanoreceptors in all experiments for feeling the surfaces they were coming into contact with. The movement towards objects during the experiments, rather than the direct path to the petrie dish edge demonstrated that either chemoreception or photoreception was being used. The percentage of individuals that went towards the coloured plastic was relatively low; however, it was shown to be much greater than the clear plastic. This demonstrates that individuals were using photoreceptors as part of its movement behaviour. A higher movement of individuals towards the plant matter, in comparison to the other objects, would suggest that chemosensory is playing a determinate role in the unknown Risoid species'. However, results were not consistent throughout the replicates. This may be because individuals were reused over a short period of time or simply because some individuals were searching for food, whilst others protection.
Functional Biology
Reproduction
Most of the Rissoidae family are gonochoric, meaning that individuals are separate sexes (Ruppert,et al. 2004). The gonad (either ovary or testies) are located in the spirals of the visceral mass. Males have a penis which sits behind the right eye in all Rissoidae species (Ponder, 1985).The female genitalia vary greatly between genera of the Rissoidae and are therefore used as a defining characteristic between some species (Ponder,1985). Majority of the Rissoid family show internal fertilisation at the female’s posterior end (Ponder, 1985). Egg capsules have been described as circular to lens shape with a thin outer covering (Ponder, 1985). Numerous surfaces have been observed for egg laying, including rock, algae and seagrass (Ponder,1985).
Some genus’s, such as Voorwindia, demonstrate development with a planktotrophic larvae, while others have been shown to use direct development (Ponder, 1985). Species that undergo direct development generally lay fewer eggs of larger size than species which have planktotrophic larvae (Ponder, 1985). This is because the species that undergo direct development have eggs with higher yolk content (Ponder, 1985).
Mini Experiment:
20 of the Rissoidae individuals were collected from the aquarium and placed in an 8cm petrie dish with some plant matter. The petrie dish was left for a week, with small water changes done on a daily basis to keep the population alive. After a week the sample was carefully observed to spot any eggs or juveniles. This was successful with eggs found at two different stages, as well as juveniles. The eggs and juveniles were found on the plant matter left in the petrie dish. These were measured and placed under compound microscopes for better detail (egg; figure 9 & juveniles; figure 10). This demonstrates that the unidentified Rissoidae species is direct developing and has a fast reproduction cycle which explains its success in colonizing areas. This is a major characteristic that seperates the unidentified species from the genus Voorwindia which produce planktotrophic larvae.
Eggs were found at a size of 0.22-0.25mm with settled juveniles from 0.3mm.
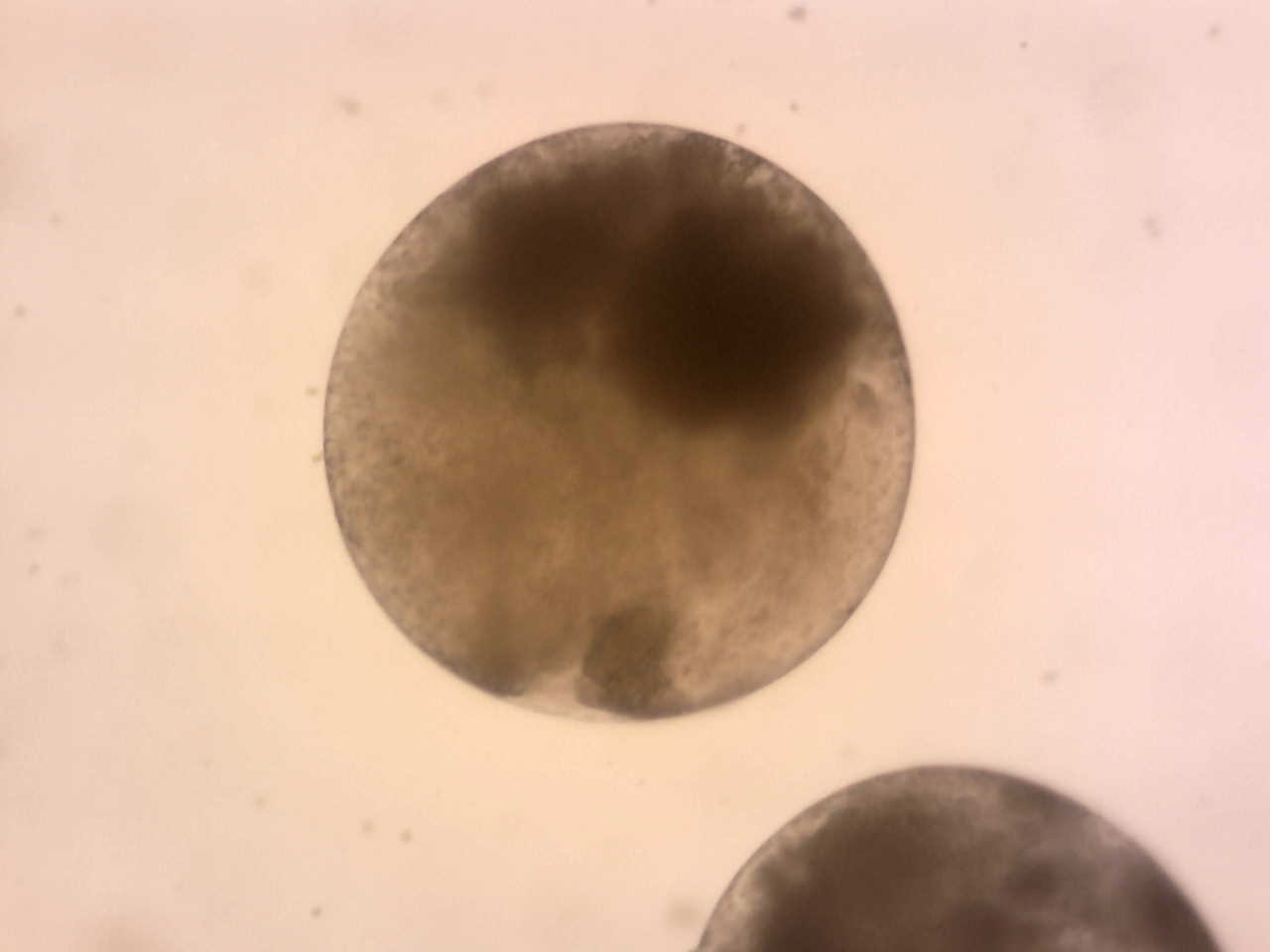 Figure 9: Shows eggs from the unidentified Rissoidae
Figure 9: Shows eggs from the unidentified Rissoidae
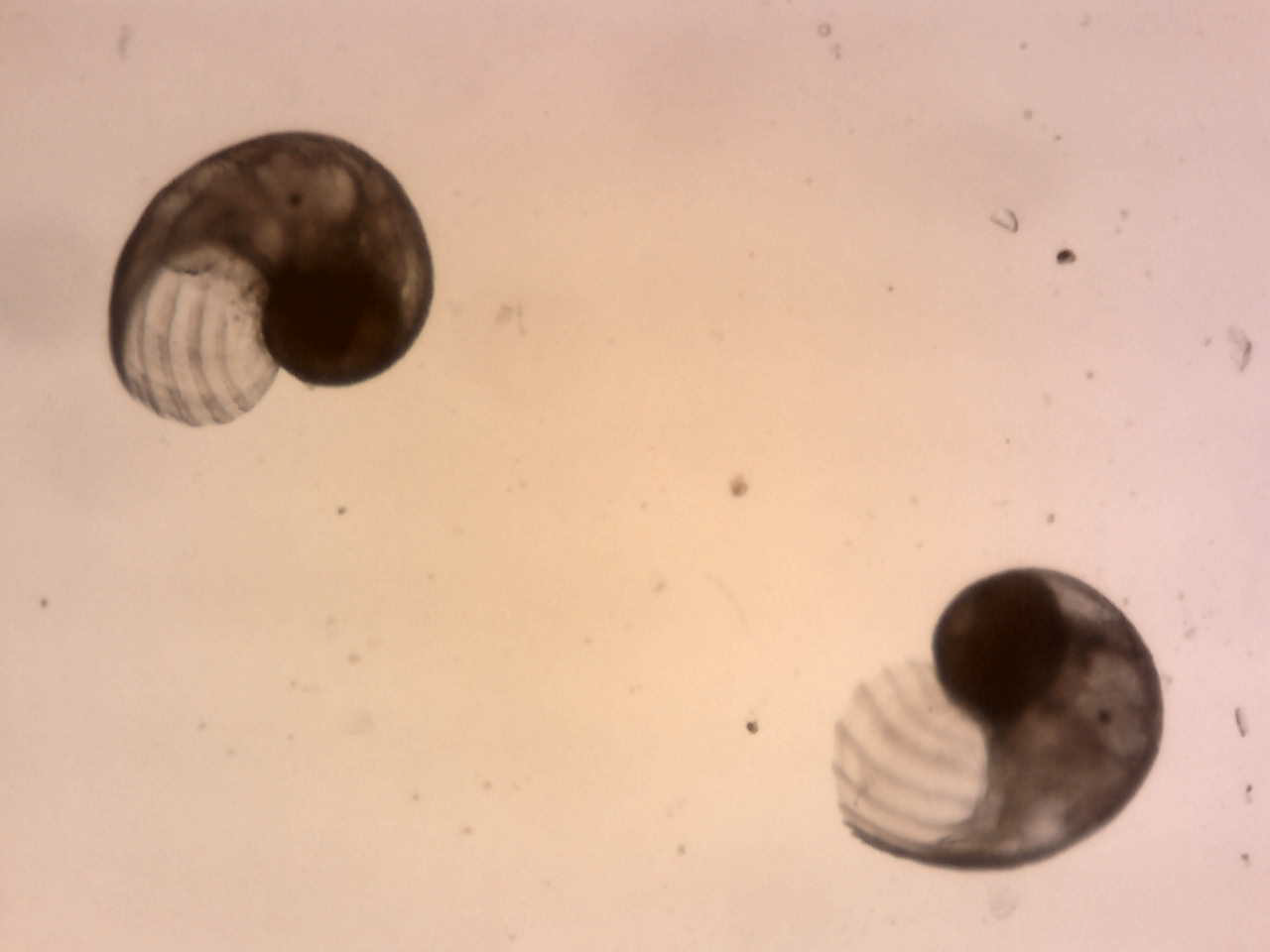 Figure 10: Shows juveniles of the unidentified Rissoidae species.
Figure 10: Shows juveniles of the unidentified Rissoidae species.
Diet:
The main diet of Rissoid members is the microalgae and diatomaceous films that live on macro algae surfaces (Ponder, 1985). This explains the shallow distribution of most species in this family. Ponder (1985) describes that some deep-sea and some shallow-water species are selective deposit feeders. A number of species have also been found to eat foraminifera; however, their stomach contents showed this was not their only food source (Ponder, 1985).
Respiration:
A generalized molluscan gill consists of a central axis, which encloses nerves, blood vessels and muscle (Ruppert,et al. 2004). From the axis two staggered rows of gill filaments arise, this is known as a bipectinate gill (Ruppert,et al. 2004).However, members of the mesogastropoda have lost their right gill, making them monopectinate (Ruppert,et al. 2004; Ponder, 1985).Cilia on the remaining gill filament creates counter current water flow past the gills, allowing for gas exchange from high oxygen content (water) to low oxygen content (Blood – hemolymph) (Ruppert,et al. 2004). From here the oxygenated blood is moved to the atrium before being distributed through the body (Ruppert,et al. 2004).The figure below is a replicate from Ruppert,et al. (2004), it demonstrates the blood flow in a mesogastropod. The figure provides a visual representation of the counter current gill respiration, with links to the rest of the circulatory system.
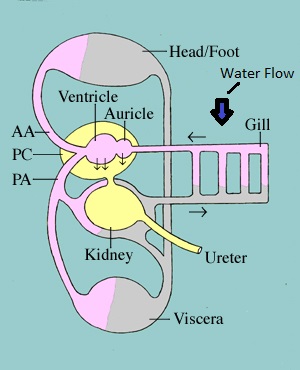 Figure 11: Demonstrates the circulatory system of a mesogastropod, showing how the gill is orientated to water current. Sourced from: The Archered shell collection of Washington State Universities Natural History Museum.
Anatomy & Physiology
A description of the morphological and physiological adaptations underlying life history traits. This may include studies at the level of cells, tissues and organs and may include dissections and the production of microscopic slides. Good photos are critical is these descriptions.
External Anatomy
The unidentified Risoid species is a small gastropod (~2.3mm) with white/opaque body colour. The shell itself is elongate-conical shell, partially transparent, smooth and dextrally whorled (~3.25 whorls). The protoconch resembles a dashed, spiralling pattern. Individuals demonstrated a well developed head and two cephalic tentacles, with a lateral eye at the base of each. The muscular foot is long and slender with posterior and anterior pedal glands present. A thin operculum was observed when the individuals retracted.
External Shell Anatomy:
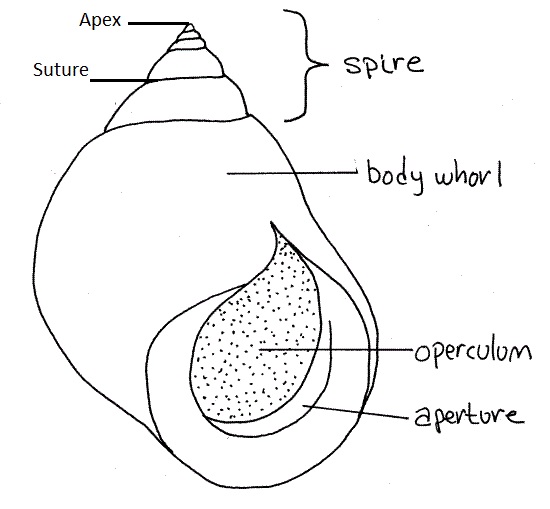
Figure 12: Was sourced from a site looking at the anatomy of Littorina irrorata, to show the shell characteristics of a mesogastropod.
Apex: The point of the shell where the whorl is smallest; also the part of the shell that developed first.
Suture: the spiralled depression on the shell which separates the whorls.
Whorl: A whorl is a 360o rotation of the shell.
Body Whorl: The largest and most recently formed whorl.
Spire: Consists of all the whorls except for the body whorl.
Aperture: The large opening of the shell, through which the head and foot can be retracted.
Operculum: A firm disk, found on the posterior dorsal surface, that when retracted seals off the shells entrance.
Note: Above information from Ruppert (2004).
Internal Anatomy
Like all gastropods, Rissoidae undergo torsion during their larval development. Ruppert, et al.(2004) explains that torsion is the 180o anticlockwise rotation of the visceral mass, shell, mantle, and mantle cavity with respect to the head and foot. Prior to torsion a gastropods organs extend down to its posterior end (Ruppert, et al. 2004). Torsion results in the movement of the mantle cavity and organs behind and directly above the head at the anterior end, as visible in figure 13 (Ruppert,et al. 2004). The stomach is positioned dorsally at the posterior, while the mouth and anus are now anterior and ventral, creating a U-shaped digestive system (Ruppert,et al. 2004).
Similarly, the nervous system is twisted into a figure-eight shape, with the visceral ganglion at the base and the nerve ring positioned at the top of the figure eight (Ruppert,et al. 2004). A full digestive system is present, with an anus and nepridia for the release of waste. Molluscs have a central nervous system that consists of several pairs of ganglia. The cerebral ganglion (brain) is connected with the eyes, statocysts and other sense organs in molluscs. A number of other ganglia pairs are also in place to innervate the muscular foot, radular muscles and mantle.
Rissoidae have one nephridium which is a blind sac, located anteriorly, and surrounded by hemocoel. This functions in the removal of metabolic waste products in the blood. Waste is excreted from the nephridopore, which in the prosobranchs is located at the mantle cavities rear, downstream from the gills.
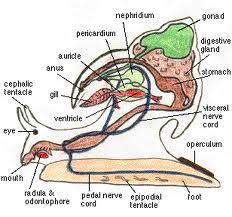
Figure 13: Mesogastropod internal anatomy sourced from a webpage looking at snail distribution in regards to temperature.
Cephalic Tentacles: sensitive projections often used for sensing water vibrations and feeling surrounding objects.
Radula: transverse rows of small chitinous teeth, part of the anteriorly positioned mouth.
Foot: Fleshy, muscular structure used for locomotion, plus hunting and reproduction in some taxa. (More information of muscular foot under Appearance).
Gill: counter current respiratory structure that may be bipectinate or monopectinate in gastropods. Mesogastropoda are Monopectinate representatives.
Visceral Nerve Cord: Nerve cord running posteriorly from the cerebral ganglion to the visceral ganglia (Rawat, 2010).
Pedal nerve cord: Nerve cords running ventrally from the cerebral ganglion to another ganglion in the foot (Rawat, 2010).
Cerebral Ganglion: concentrations of nerve tissue situated above the digestive tract. Gives rise to nerves that provide the head region and associated sense organs. May connect additional ganglion to form central nervous system (circumenteric nerve ring).
Stomach: weakly muscularized region of the digestive tract between the oesophagus and intestine. Important location of food digestion.
Digestive gland: sometimes referred to as the midgut gland, this structure specialises in the creation of digestive fluids.
Gonad: may be the ovaries or testies depending on if it is male or female specimen.
Nephridium: Release metabolic waste products from the blood into the mantle cavity, waste removed by the current flowing out of the shell once past the gills.
Pericardium: a thin walled, ovoid chamber that encircles the heart.
Auricle: Part of an open circulatory system, collects oxygen rich blood from the gills.
Ventricle: Pumps blood from the auricle into small vesicles, carrying the oxygen enriched blood to the organism’s tissues.
Note: Above information from Ruppert, et al. (2004).
Evolution & Systematics
Using existing literature, describe what is known about the systematics of the target species/genus, working from the phylum down to the lowest taxonomic unit. Where possible describe what is known about the evolution of this group, drawing upon phylogenetic, paleontological and other research.
Fossil History
The fossil history for Rissoidae is relatively poor, probably due to the hard, shallow-water environments that most members live in(Ponder, 1985). The Rissoids were first known to appear during the Middle Jurassic, with remains found in Europe. The fossil history was relatively poor during the Cretaceous but showed high diversities during the Tertiary. Many of the extant taxa have fossil records dating back to the Eocene (Ponder, 1985).
Phylogenetics
As mentioned, the family Rissoidae supports a global distribution of small, marine and brackish species. Classifying members of the Rissoid family, based on morphological characteristics, has proven to be very difficult. The diverse form, small size and convergence of characteristics have proven to be the trickiest obstacles in Rissoidae classification. The gastropod at the focus of this study shows the morphological characteristics of the family Rissoidae. On the genus level it becomes much more difficult with similarities to the genera Voorwindia and Setia.
The main issue with classifying the unidentified Rissoidae to Voorwindia is the development processes. The unidentified specimen demonstrates direct development; whilst Ponder (1985) explains that all Voorwindia have planktotrophic larvae. Setia on the other hand do show direct development but have not been sampled near Australia (Avila, 2013). This does not mean they aren't present but is a limiting factor when trying to determine the unidentified gastropods genus. Voorwindia on the other hand have been sampled in numerous areas of Australia's east coast, which would make them a good candidate if it wasn't for its development characteristics (Ponder, 1985).
Kingdom: Animalia
Phylum: Mollusca
Class: Gastropoda
Order: Littorinimorpha
Sub-Order: Mesogastropoda
Superfamily: Risooidea
Family: Risoidae
genus: Unknown (Perhaps Voorwindia or Setia)
Species: Unknown
Threats
Invasion of environments by other organisms has the ability to cause major population fluctuations in any ecosystem (Alonso &Castro-Diez, 2012). In the case of Rissoidae, their small size could make them easy prey for invading organisms. Competition for food against similar gastropods, or other marine organisms, could also cause increased population pressures on a species (Alonso & Castro-Diez, 2012).
Another major threat to Rissoidae species may come in the form of anthropogenic activities. A paper by Smith (2006) investigates the affects eight chemicals, which are often used on yachts, have on the gastropod Nassarius obsoletus; this was undertaken after members of the species showed sexual abnormities. High coastal populations have been shown to cause major changes in the chemical, physical and biological properties of estuaries. This may come in many forms, including the release of pollutants into the estuary, replacement of natural shorelines with hard surfaces and the alteration of water flow (Inglis & Cross, 2000).Such significant changes have been shown to alter the invertebrate communities which inhabit these ecosystems (Inglis & Cross, 2000).
Global warming has been observed over the past century with evident ties to anthropogenic activity (Doney, et al. 2012). The warming of earth’s oceans and rising sea levels may put pressure on species to relocate their distribution on a local and regional scale (Doney, et al. 2012). For species that are dependent on micro/macro algae, rises in sea-level has the potential to put major stresses on both the producer and consumer (Doney, et al. 2012). Earth’s oceans absorb a large quantity of the atmospheric CO2; this has increased as greenhouse gas emissions have (Hendricks, et al. 2010). As a result, there have been recordings of change in ocean pH, with a trend of acidification (Hendricks, et al. 2010). Ocean acidification has been a proposed threat to invertebrates that are shell-forming or calcareous (Hendricks, et al. 2010). However, it is hard to determine whether these organisms will begin to die-off or adapt to the changing conditions.
References & Links
Alonso, A., & Castro-Diez, P. (2008). What explains the invading success of the aquatic mud snail Potamopyrgus antipodarum (Hydrobiidae, Mollusca)?.Hydrobiologia, 614(1), 107-116.
Ávila, S. P. (2013). Unravelling the patterns and processes of evolution of marine life in oceanic islands: a global framework. Climate Change perspectives from the Atlantic: past, present and future. Tenerife, Universidad de La Laguna, 85-115.
Doney,SC, Ruckelshaus, M, Duffy, JE, Barry, JP, Chan, F, English, CA, Galindo, HM, Grebmeier, JM, Hollowed, AB,Knowlton, N, Polovina, J, Rabalais, NN, Sydeman, WJ & Talley, LD 2012.‘Climate change impacts on marine ecosystems’, Marine Science, vol. 4, pp. 11-47.
Hendriks, I. E., Duarte, C. M., & Álvarez, M. (2010).Vulnerability of marine biodiversity to ocean acidification: a meta-analysis. Estuarine, Coastal and Shelf Science, 86(2), 157-164.
Inglis, G. J., & Kross, J. E. (2000). Evidence for systemic changes in the benthic fauna of tropical estuaries as a result of urbanization. Marine Pollution Bulletin, 41(7), 367-376.
Ponder, W. F. (1985). A review of the genera of the Rissoidae (Mollusca: Mesogastropoda: Rissoacea). Australian Museum.
Rawat, R. (2010). Anatomy of Mollusca. International scientific publishing academy. New Dehli, India.
Ruppert, E. E., Barnes, R. D., & Fox, R. S. (2004). Invertebrate zoology. Brooks/Cole, Cengage Learning.
Smith, B. S. (1981). Male characteristics on female mud snails caused by antifouling bottom paints. Journal of Applied Toxicology, 1(1), 22-25.
|